Клеточная
мембрана
У этого термина существуют и другие
значения, см. Мембрана.
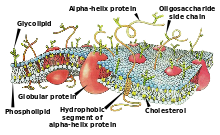
Изображение клеточной мембраны. Маленькие
голубые и белые шарики соответствуют гидрофильным
«головкам» фосфолипидов, а присоединённые к ним линии — гидрофобным «хвостам».
На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые
спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Жёлто-зеленые
цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов,
формирующие гликокаликс.
Кле́точная мембра́на (также цитолемма, плазмалемма, или плазматическая мембрана) —
эластическая молекулярная структура, состоящая из белков и липидов. Отделяет содержимое
любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между
клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные
замкнутые отсеки —
компартменты
или органеллы,
в которых поддерживаются определённые условия среды.
Основные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных
клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой
двойной слой (бислой)
молекул класса липидов,
большинство из которых представляет собой так называемые сложные липиды — фосфолипиды.
Молекулы липидов имеют гидрофильную
(«головка») и гидрофобную
(«хвост») части. При образовании мембран гидрофобные участки молекул оказываются
обращены внутрь, а гидрофильные —
наружу. Мембраны — структуры инвариабельные,
весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи,
у которых мембраны образованы глицерином
и терпеноидными
спиртами. Толщина мембраны составляет 7—8 нм.
Биологическая мембрана включает и различные
белки:
интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним
концом во внешний или внутренний липидный слой), поверхностные (расположенные на
внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются
точками контакта клеточной мембраны с цитоскелетом
внутри клетки, и клеточной
стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют
функцию ионных каналов,
различных транспортеров
и рецепторов.
История исследования
В 1925 году Гортер и Грендель с помощью
осмотического удара получили так называемые «тени» эритроцитов —
их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем
с помощью ацетона выделили из оболочек липиды и определили количество липидов на
единицу площади эритроцита — этого количества хватило на сплошной двойной слой.
Хотя этот эксперимент привел исследователей к правильному выводу, ими было допущено
несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно
все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому
весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей
случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными
пленками показали, что они обладают высоким поверхностным натяжением, гораздо большим,
чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение —
белки. В 1935 году Даниэлли и Доусон представили научному сообществу модель «сендвича»,
которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам
от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические
исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были
видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один
электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал
в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трехслойное
строение всех клеточных мембран.
Но постепенно накапливались аргументы
против «бутербродной модели»:
• накапливались сведения о глобулярности плазматической
мембраны
• оказалось, что структура мембраны при электронной
микроскопии зависит от способа её фиксации
• плазматическая мембрана может различаться
по структуре даже в одной клетке, например в головке, шейке и хвосте сперматозоида
• «бутербродная» модель термодинамически не
выгодна — для поддержания такой структуры нужно затрачивать большое количество
энергии, и протащить вещество через мембрану очень сложно
• количество белков, связанных с мембраной
электростатически очень небольшое, в основном белки очень тяжело выделить из мембраны,
так как они погружены в неё
Все это привело к созданию в 1972 году
С. Д. Сингером (S. Jonathan Singer) и Г. Л. Николсоном
(Garth L. Nicolson) жидкостно-мозаичной модели строения мембраны. Согласно этой
модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные,
полуинтегральные и периферические. Периферические белки действительно находятся
на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими
взаимодействиями, но никогда не образуют сплошной слой. Доказательствами жидкостности
мембраны служат методы FRAP,
FLIP и
соматическая
гибридизация клеток, мозаичности — метод
замораживания-скалывания, при котором на сколе мембраны видны бугорки
и ямки, так как белки не расщепляются, а целиком отходят в один из слоев мембраны.
Функции
• = барьерная — обеспечивает регулируемый, избирательный, пассивный
и активный обмен веществ с окружающей средой. Например, мембрана пероксисом
защищает цитоплазму от опасных для клетки пероксидов.
Избирательная
проницаемость означает, что проницаемость мембраны для различных атомов
или молекул
зависит от их размеров, электрического
заряда и химических свойств. Избирательная проницаемость обеспечивает
отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми
веществами.
• = транспортная — через мембрану происходит транспорт веществ в клетку
и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ,
удаление конечных продуктов обмена, секрецию различных веществ, создание ионных
градиентов, поддержание в клетке оптимального pH и концентрации
ионов, которые нужны для работы клеточных ферментов.
• Частицы, по какой-либо причине неспособные
пересечь фосфолипидный бислой
(например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает
гидрофильные
вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть
сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы
или путём эндоцитоза.
• При пассивном
транспорте вещества пересекают липидный бислой
без затрат энергии по градиенту концентрации (градиент концентрации указывает направление
увеличения концентрации) путём диффузии. Вариантом этого механизма является облегчённая
диффузия,
при которой веществу помогает пройти через мембрану какая-либо специфическая молекула.
У этой молекулы может быть канал, пропускающий вещества только одного типа.
• Активный
транспорт требует затрат энергии, так как происходит против градиента
концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза,
которая активно вкачивает в клетку ионы калия
(K+) и выкачивает из неё ионы натрия
(Na+).
• = матричная — обеспечивает определенное взаиморасположение и
ориентацию мембранных белков, их оптимальное взаимодействие.
• = механическая — обеспечивает автономность клетки, её внутриклеточных
структур, также соединение с другими клетками (в тканях). Большую роль в обеспечении
механической функции имеют клеточные стенки,
а у животных — межклеточное
вещество.
• = энергетическая — при фотосинтезе
в хлоропластах
и клеточном
дыхании в митохондриях
в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
• = рецепторная — некоторые белки, находящиеся в мембране, являются
рецепторами
(молекулами, при помощи которых клетка воспринимает те или иные сигналы).
• Например, гормоны,
циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие
этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение
нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
• = ферментативная — мембранные белки нередко являются
ферментами.
Например, плазматические мембраны эпителиальных
клеток кишечника содержат пищеварительные ферменты.
• = осуществление генерации и проведения биопотенциалов:
• С помощью мембраны в клетке поддерживается
постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно
выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно,
так как это обеспечивает поддержание разности потенциалов на мембране и генерацию
нервного
импульса.
• = маркировка клетки — на мембране есть антигены,
действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины
(то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми
цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых
цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров
клетки могут распознавать другие клетки и действовать согласованно с ними, например,
при формировании органов и тканей. Это же позволяет иммунной системе
распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов:
фосфолипиды,
гликолипиды
и холестерол.
Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из
двух длинных гидрофобных углеводородных
«хвостов», которые связаны с заряженной гидрофильной
«головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство
между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны
с малым содержанием холестерола более гибкие, а с большим — более жёсткие и
хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных
молекул из клетки и в клетку.
Важную часть мембраны составляют белки,
пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация
в разных мембранах различаются. Рядом с белками находятся аннулярные липиды —
они более упорядочены, менее подвижны, имеют в составе более насыщенные жирные кислоты
и выделяются из мембраны вместе с белком. Без аннулярных липидов белки мембраны
не работают.
Клеточные мембраны часто асимметричны,
то есть слои отличаются по составу липидов, в наружном содержатся преимущественно
фосфатидилинозитол,
фосфатидилхолин,
сфингомиелины
и гликолипиды,
во внутреннем — фосфатидилсерин,
фосфатидилэтаноламин
и фосфатидилинозитол.
Переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп)
затруднён, но может происходить спонтанно, примерно раз в 6 месяцев или с помощью
белков-флиппаз
и скрамблазы
плазматической мембраны. Если в наружном слое появляется фосфатидилсерин, это является
сигналом для макрофагов
о необходимости уничтожения клетки.
Мембранные органеллы
Это замкнутые одиночные или связанные
друг с другом участки цитоплазмы,
отделённые от гиалоплазмы
мембранами. К одномембранным органеллам относятся эндоплазматическая
сеть, аппарат Гольджи,
лизосомы,
вакуоли,
пероксисомы;
к двумембранным — ядро,
митохондрии,
пластиды.
Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной
проницаемостью: через них медленно диффундируют глюкоза,
аминокислоты,
жирные кислоты,
глицерол
и ионы,
причем сами мембраны в известной мере активно регулируют этот процесс — одни
вещества пропускают, а другие нет. Существует четыре основных механизма для поступления
веществ в клетку или вывода их из клетки наружу: диффузия,
осмос,
активный
транспорт и экзо-
или эндоцитоз.
Два первых процесса носят пассивный характер, то есть не требуют затрат энергии;
два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны
при пассивном транспорте обусловлена специальными каналами — интегральными
белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для
элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы
этих элементов движутся в клетку и из неё. При раздражении каналы натриевых
ионов раскрываются, и происходит резкое поступление в клетку ионов натрия.
При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал
восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают
ионы калия.
См. также
• Липиды
Литература
• Антонов В. Ф., Смирнова Е. Н., Шевченко Е.
В. Липидные мембраны при
фазовых переходах. — М.: Наука, 1994.
• Геннис Р. Биомембраны. Молекулярная структура и функции: перевод
с англ.
= Biomembranes. Molecular structure and function (by Robert B. Gennis). —
1-е издание. — М.: Мир, 1997. — ISBN
5-03-002419-0.
• Иванов В. Г., Берестовский Т. Н. Липидный бислой
биологических мембран. — М.: Наука, 1982.
• Рубин А. Б. Биофизика,
учебник в 2 тт. — 3-е издание, исправленное и дополненное. —
М.: издательство Московского университета, 2004. — ISBN
5-211-06109-8.
• Bruce Alberts, et al. Molecular
Biology Of The Cell. — 5th ed. — New York: Garland
Science, 2007. — ISBN
0-8153-3218-1. — учебник по молекулярной биологии на английском
языке
Ссылки
• Владимиров Ю.
А., Повреждение компонентов биологических мембран при патологических процессах
Категории: Цитология Клеточная
мембрана Органеллы
Источник: Википедия
 Главная
Главная Альтернативная медицина
Альтернативная медицина